Accueil / Accueil – articles / La Sardaigne préhistorique





La Sardaigne préhistorique
La Sardaigne préhistorique
Origine et peuplement préhistorique
La Sardaigne et la Corse sont deux des plus grandes îles de la Méditerranée. Elles étaient reliées entre elles au dernier maximum glaciaire (LGM). Ceci était dû au niveau de la mer qui était à cette période plus bas de 130 mètres jusqu’à il y 14 000 à 15 000 ans environ. Formant ainsi le bloc cyrno-sarde, masse continentale d’environ 35 000 km². Ce bloc formait la plus grande île de Méditerranée où affleurent les plus anciennes roches du Paléozoïque, c’est-à-dire appartenant à la chaîne varisque. Malgré sa proximité avec la Corse, la Sardaigne est non seulement très différente en topographie, mais aussi inhabituelle parmi les îles méditerranéennes en raison d’un relief relativement faible pour sa taille (24 090 km²). Cette différence serait d’origine géologique, car la Sardaigne est un mélange de types de roches comprenant des granites mais aussi du basalte, des schistes, du grès et du calcaire dolomitique. La présence en Sardaigne de précieuses ressources minérales, notamment l’obsidienne du Mont-Arci, le cuivre, l’étain, le fer et le plomb aurait entraîné l’occupation humaine et le changement du paysage, en particulier dans les basses terres (voir Brown & Walsh, 2016).
Un primate en Sardaigne ?
Un primate tout à fait spécial fréquentait cette île, il s’agissait d’un hominoïde du Miocène, l’espèce Oreopithecus bambolii, qui signifie « singe des collines de Bamboli ». Ce primate insulaire de la Toscane actuelle et de la Sardaigne (site de Fiume Santo) est à ce jour le dernier hominoïde enregistré d’Europe (-8,3 à -6,7 Ma) et l’un des rares connus à avoir évolué dans des conditions insulaires. Une étude récente indique qu’il n’utilisait pas la bipédie permanente (il aurait probablement pu se déplacer en bipédie à certains moments) car, des détails de l’anatomie montrent qu’il diffère à la fois des grands singes existants (chimpanzés, gorilles…) et des premiers hominidés (Ardipithecus ramidus, Australopithecus…) et ce, de manière clé. Cette étude clarifie les scénarios phylogénétiques et évolutifs de cette espèce en 3 possibilités à ce jour. En résumé, soit Oreopithecus pourrait être un membre de la tige des Hominidae, soit de la tige des Hylobatidae ou bien finalement être le résultat d’une convergence évolutive et donc aligné avec n’importe quel groupe hominoïde (p. ex., les hominoïdes souches, les hylobatidés ou les hominidés ; voir Hammond et coll., 2020).
Peuplements humains de l’île

Les restes humains les plus anciens connus en Sardaigne remonteraient au moins à 20 000 ans environ et ont été découverts dans la grotte Corbeddu (Oliena). Des preuves archéologiques suggèrent que la Sardaigne n’était pas densément peuplée au Mésolithique et a connu une expansion démographique coïncidant avec la transition néolithique au VIème millénaire AEC. À cette époque, les premiers assemblages de poterie néolithique se répandaient dans toute la Méditerranée occidentale, y compris la Sardaigne, en particulier les récipients décorés d’empreintes de coquille de cardium, avec des dates de radiocarbone indiquant une expansion maritime rapide vers l’ouest vers 5 500 AEC. Au Néolithique tardif, l’obsidienne originaire de Sardaigne se retrouve dans de nombreux sites archéologiques de la Méditerranée occidentale, ce qui indique que l’île a été intégrée dans un réseau de commerce maritime. Au milieu de l’âge du Bronze, vers 1 600 AEC, la culture nuragique a émergé, du nom des milliers de tours de pierre distinctives, appelées nuraghes.
À la fin de la période nuragique, les archives archéologiques et historiques montrent l’influence directe de plusieurs grands groupes méditerranéens, en particulier la présence de commerçants mycéniens, levantins et chypriotes. Les colonies nuragiques ont décliné dans une grande partie de l’île car, à la fin du 9ème et au début du 8ème siècle AEC, les Phéniciens originaires du Liban actuel et du nord de la région géographique palestinienne, ont établi des colonies concentrées le long des rives sud de la Sardaigne. Dans la seconde moitié du 6ème siècle AEC, l’île était occupée par des Carthaginois (également connus sous le nom de Puniques), s’étendant de la ville de Carthage sur la côte nord-africaine de la Tunisie actuelle, fondée à la fin du 9ème siècle par les Phéniciens. La Sardaigne a été occupée par les forces romaines en 237 AEC et est devenue une province romaine une décennie plus tard. Tout au long de la période impériale romaine, l’île est restée étroitement alignée avec l’Italie et le centre de l’Afrique du Nord. En 170 EC, Pausanias qualifia le climat de Sardaigne de « malsain et pestilentiel », et l’empereur Néron bannit les ennemis politiques dans l’espoir qu’ils meurent du paludisme (la maladie a continué d’être un problème majeur pour le développement humain et l’expansion sur l’île jusqu’en 1951). Après la chute de l’empire romain, la Sardaigne est devenue de plus en plus autonome, mais l’interaction avec l’Empire byzantin, les républiques maritimes de Gênes et de Pise, le royaume catalan et aragonais et le duché de Savoie et du Piémont avait continué d’influencer l’île. (voir Marcus et coll. 2020).
Anthropologie génétique
S’il y a bien un sujet dans le coeur des sardes, c’est celui de leurs origines. La génétique des populations de la Sardaigne est étudiée depuis longtemps, en partie en raison de son importance pour la génétique médicale. La Sardaigne possède un patrimoine archéologique, linguistique et culturel unique et fait partie des réseaux commerciaux méditerranéens depuis le Néolithique. On ignore cependant dans quelle mesure l’ascendance génétique de la population a changé au cours de cette période. Pour générer une nouvelle perspective sur l’histoire génétique de la Sardaigne, un groupe interdisciplinaire composé de généticiens, d’archéologues et d’anciens experts en ADN ont publié cette année un article (Marcus et coll. 2020) concernant le séquençage de l’ADN ancien de 70 individus provenant de 21 sites archéologiques sardes couvrant le Néolithique moyen jusqu’à la période médiévale. L’échantillonnage d’ADN à partir de restes humains anciens permet aux scientifiques d’obtenir un certain instantané des personnes vivant à un moment et à un endroit spécifique. Lorsque l’équipe a comparé l’ADN de 70 individus anciens collectés en Sardaigne à l’ADN d’autres individus, ils ont découvert deux modèles majeurs. Leurs résultats indiquent une connectivité génétique changeante à travers la Méditerranée sur 6 000 ans et la découverte de deux schémas majeurs.
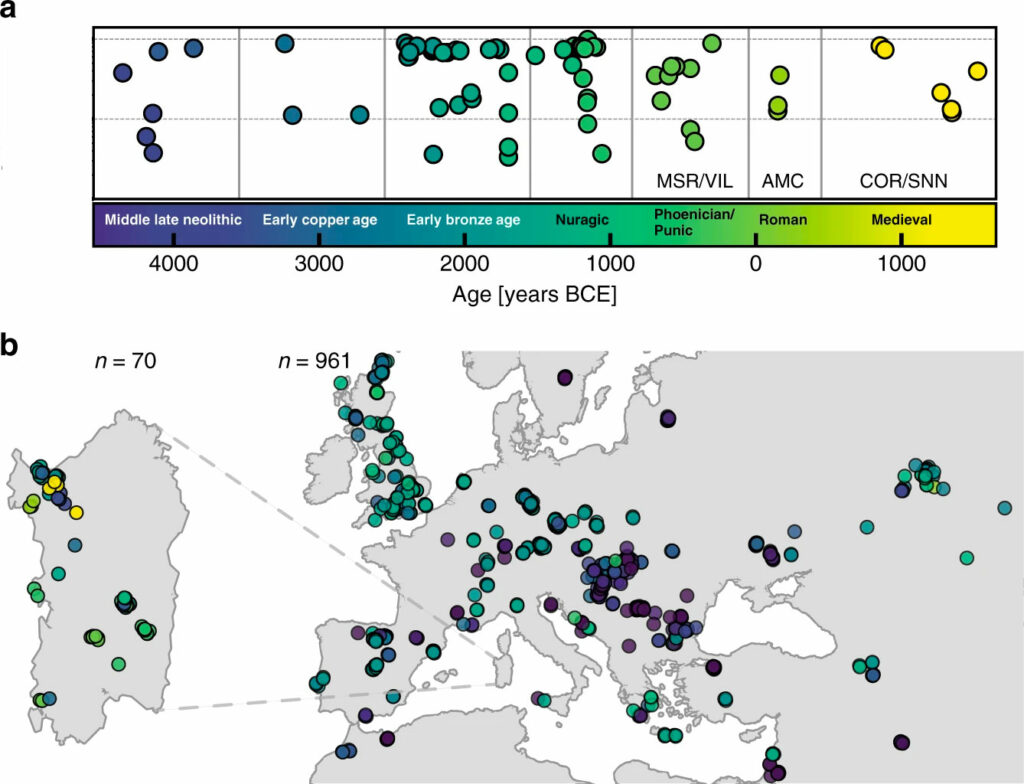
D’une part, les individus sardes du Néolithique moyen (4 100 à 3 500 AEC) étaient étroitement liés aux gens de l’Europe continentale de l’époque. L’ascendance génétique est alors restée relativement stable sur l’île au moins jusqu’à la fin de la période nuragique (environ 900 AEC). Ce modèle diffère des autres régions d’Europe continentale qui ont connu de nouveaux ancêtres provenant de personnes se déplaçant à travers le continent à l’Âge du bronze. Les résultats montrent également que le développement des nuraghes distinctives de la Sardaigne et de la culture (après laquelle la période nuragique est nommée) n’a coïncidé avec aucune nouvelle ascendance génétique détectable arrivant sur l’île. Il y aurait donc une stabilité frappante dans l’ascendance du Néolithique moyen jusqu’à la fin de la période nuragique en Sardaigne.

D’autre part, les chercheurs ont trouvé des preuves de l’arrivée de différentes populations à travers la Méditerranée, d’abord avec des Phéniciens originaires du Levant (Liban actuel) et des puniques, dont la culture était centrée à Carthage (Tunisie actuelle). Ensuite, de nouveaux ancêtres ont continué d’apparaître pendant la période romaine et plus loin pendant la période médiévale, alors que la Sardaigne était historiquement influencée par la migration de personnes de l’Italie et de l’Espagne modernes.
Il y aurait donc des signaux clairs de périodes de contacts dynamiques reliant l’île au reste de la Méditerranée, apparaissant d’abord chez des individus de deux sites phéniciens et puniques dès 500 AEC, puis chez des individus des périodes romaines et médiévales.
Ces résultats aident à expliquer les similitudes avec l’ADN d’individus d’Europe continentale du Néolithique et de l’âge du Cuivre, tel Ötzi, un humain de 5 300 ans presque parfaitement préservé découvert dans la glace alpine à la frontière italo-autrichienne en 1991. Plus précisément, parmi les Européens modernes, l’ADN d’Ötzi est le plus similaire aux Sardes modernes. La nouvelle étude soutient l’hypothèse selon laquelle cette similitude demeure parce que la Sardaigne avait moins de renouvellement de l’ascendance génétique au fil du temps que l’Europe continentale, qui a connu des migrations à grande échelle à l’Âge du bronze.
Clactonien ou Acheuléen ?

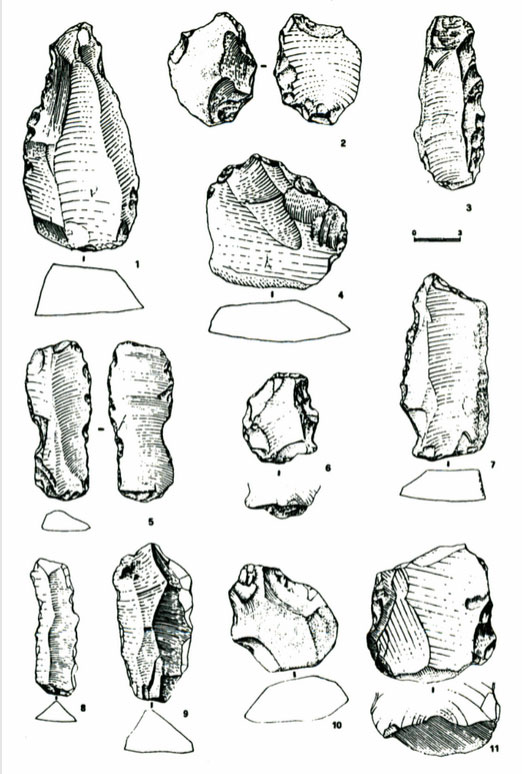
Une fréquentation de l’île par les humains semblerait présente grâce à la découverte d’industries clactoniennes localisées sur les terrasses du torrent Riu Altana, près de Perfugas (Sassari). Aussi, près de Sa Coa de Sa Multa, se trouvait un atelier de taille en place sur une haute terrasse comprenant surtout des outils de moyennes et grandes dimensions, à retouches surélevées. L’industrie roulée recueillie sur les rives du Riu Altana est un peu différente : sa laminarité est plus élevée (22 %), bien que les éclats soient dominants ; les talons sont peu inclinés et les racloirs courts à retouches sommaires, et plus rarement à retouche scalariforme ou écaillée, sont accompagnés par les denticulés, par quelques lames retouchées et un seul chopper. En général, le Clactonien de Sardaigne est de moyennes et grandes dimensions et se compose de racloirs simples et transversaux, à retouche souvent sommaire et surélevée et de denticulés ; les choppers sont rares et les bifaces sont absents. Malheureusement, le Clactonien péninsulaire n’a jamais été retrouvé en place.
L’absence totale de bifaciaux dans le Clactonien de la Sardaigne et de la Sicile semble renforcer l’hypothèse selon laquelle les complexes sur éclats seraient l’expression d’une tradition culturelle différente et autonome par rapport à l’Acheuléen (voir Grifoni & Tozzi, 2006).
Grotte de Corbeddu

À ce jour, le seul site en Sardaigne qui documente l’occupation humaine entre la glaciation du Riss et le Néolithique est la grotte de Corbeddu. À l’origine, les fouilles de cette grotte entreprises par Paul Y. Sondaar à partir de 1982 étaient concentrées sur les restes fossiles d’un cerf de Caziot (Praemegaceros cazioti), une espèce de cervidé éteinte. Mais très vite des signes de présence humaine associés à cette faune ont été trouvés dans les sédiments de la grotte à la grande surprise des archéologues. Jusque-là, la Sardaigne était considérée comme inhabitée avant le début du Néolithique. C’est le seul site archéologique connu du dernier Paléolithique glaciaire de l’île et la grotte de Corbeddu est devenue, avec la thèse de Gerard K. Hofmeijer, le site le mieux documenté du Paléolithique en Sardaigne.

La grotte de Corbeddu, formée de calcaire jurassique, est située dans la vallée de Lanaittu (Oliena). Elle doit son nom au célèbre bandit du XIXème siècle, Giovanni Corbeddu Salis, qui l’avait choisie comme refuge. Cette grotte relativement petite est formée dans le calcaire jurassique qui appartient au système Sa Oche e Su Bentu. Il s’agit d’une cavité d’aménagement presque horizontale, d’environ 130 m de long. Elle est divisée en quatre salles où des études stratigraphiques et sédimentologiques approfondies ont surtout été menées dans les salles 1 et 2. Excluant donc le lieu qu’occupait Giovanni Corbeddu Salis (salle 3) et la salle du fond qui ne servit que de fosse d’essai (Hall 4). Les fouilles de la grotte de Corbeddu étaient conformes aux normes archéologiques, en utilisant des systèmes de quadrants, en lavant les sédiments et en utilisant uniquement des outils fins et des brosses.
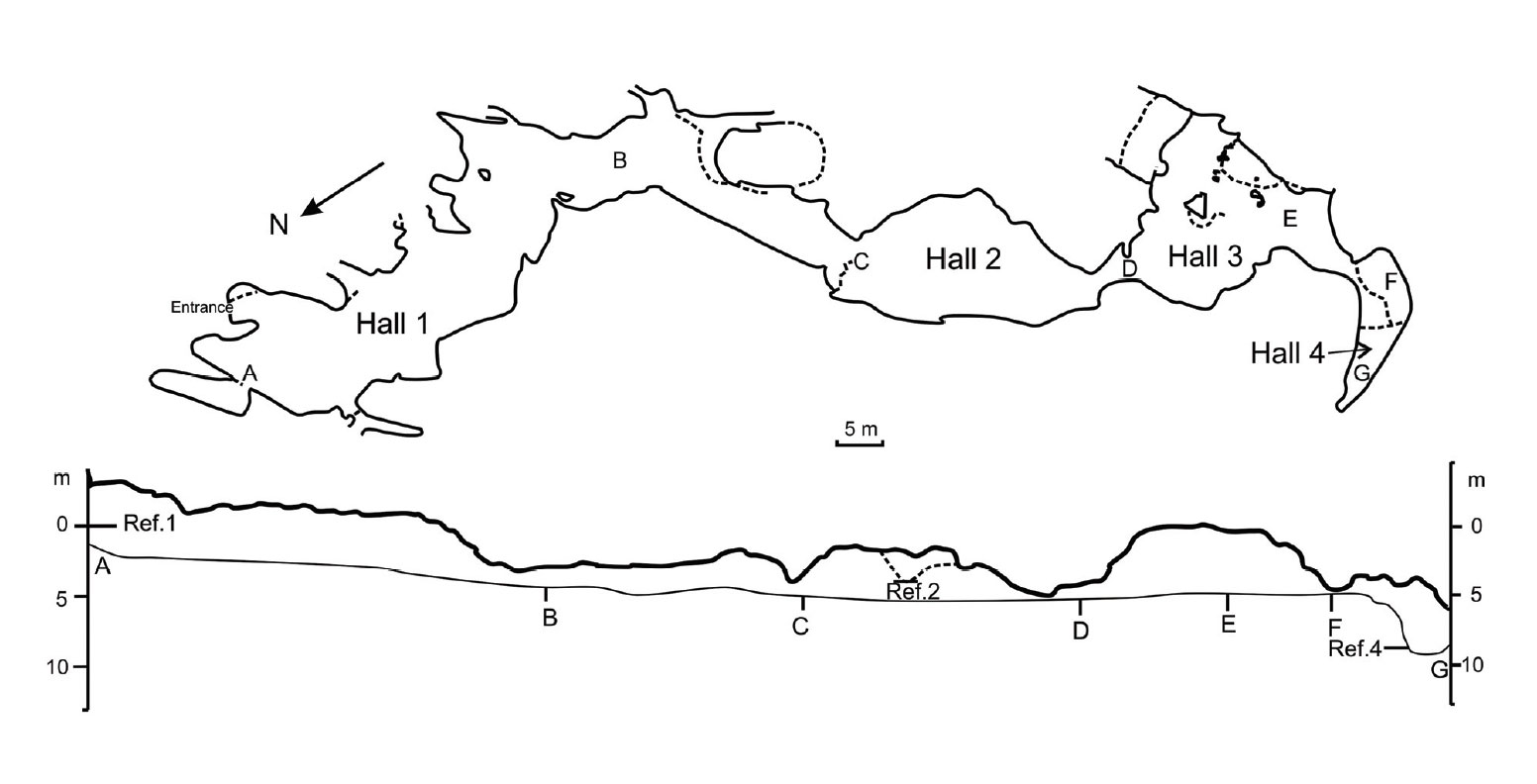
Périodiquement, probablement après des périodes de pluies abondantes, l’eau peut entrer dans la grotte de Corbeddu. Les fosses d’excavation ont été fortement endommagées en 1988 par l’eau et des travaux de réparation avaient permis la reprise des fouilles. Malheureusement, la crue de 1999 a marqué la fin des plus de quinze années de fouilles estivales. Il semble que des inondations périodiques détruisent régulièrement ce site et finiront par remplir la grotte uniformément de sédiments argileux et de brèches si des travaux de réparation n’ont pas lieu régulièrement. Cela signifie aussi que ce que l’on voit aujourd’hui dans les fosses d’excavation n’est plus représentatif des fosses du passé.
Ossements humains
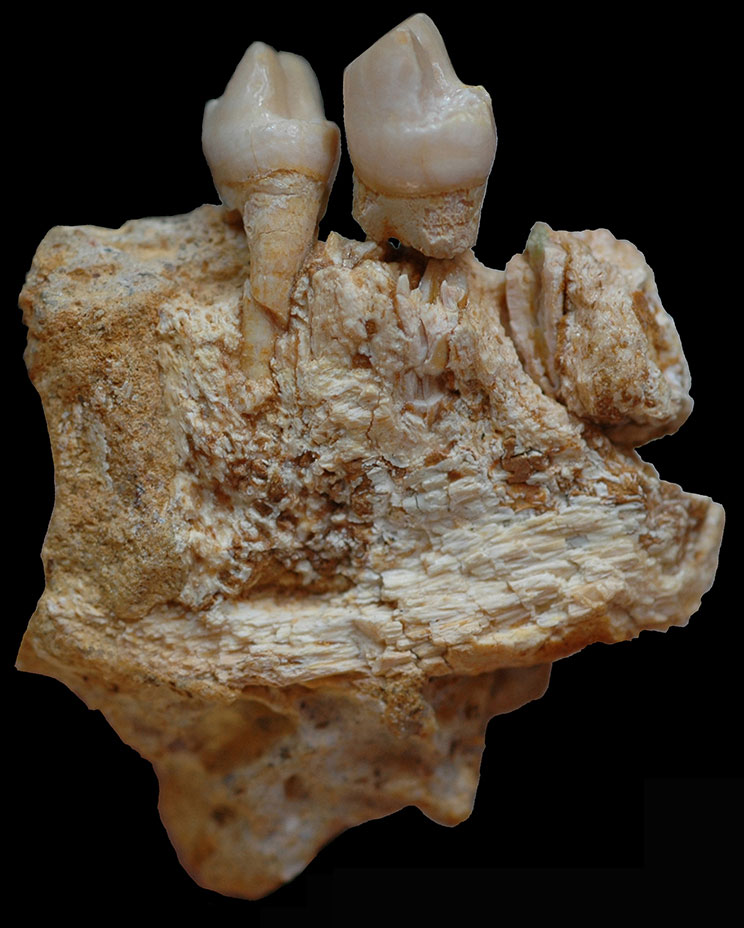
Les fouilles archéologiques dans la grotte de Corbeddu ont permis la découverte d’ossements témoignant de la présence humaine en Sardaigne au moins au Paléolithique supérieur. Une première découverte de deux fossiles humains, un os temporal droit et un maxillaire gauche (salle 2) qui seraient datés d’environ 8 750 ans BP (14C), puis quelques années plus tard, la découverte d’un fragment de phalange de main (salle 2) datant indirectement de 20 000 ans BP environ (14C et palynologie). Cela implique que ce fossile pourrait être à ce jour le plus ancien reste humain trouvé dans un contexte archéologique insulaire de la Méditerranée.
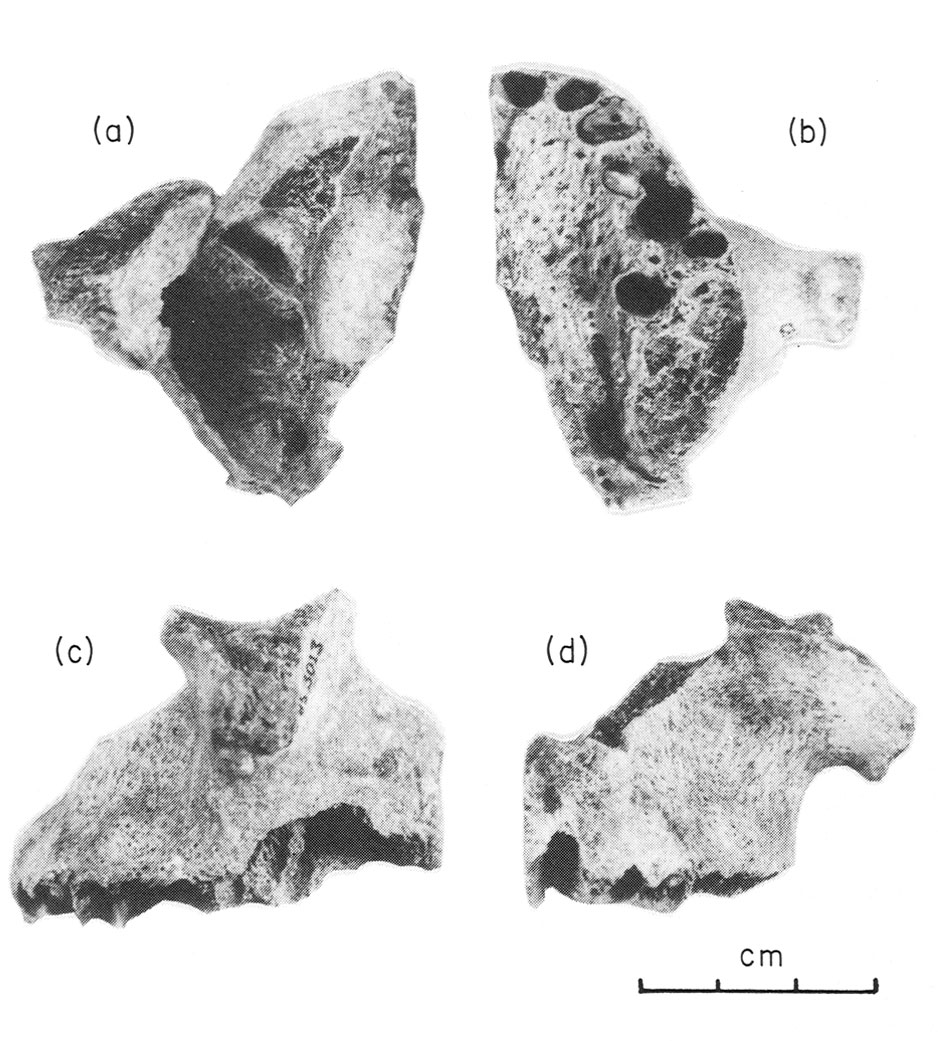
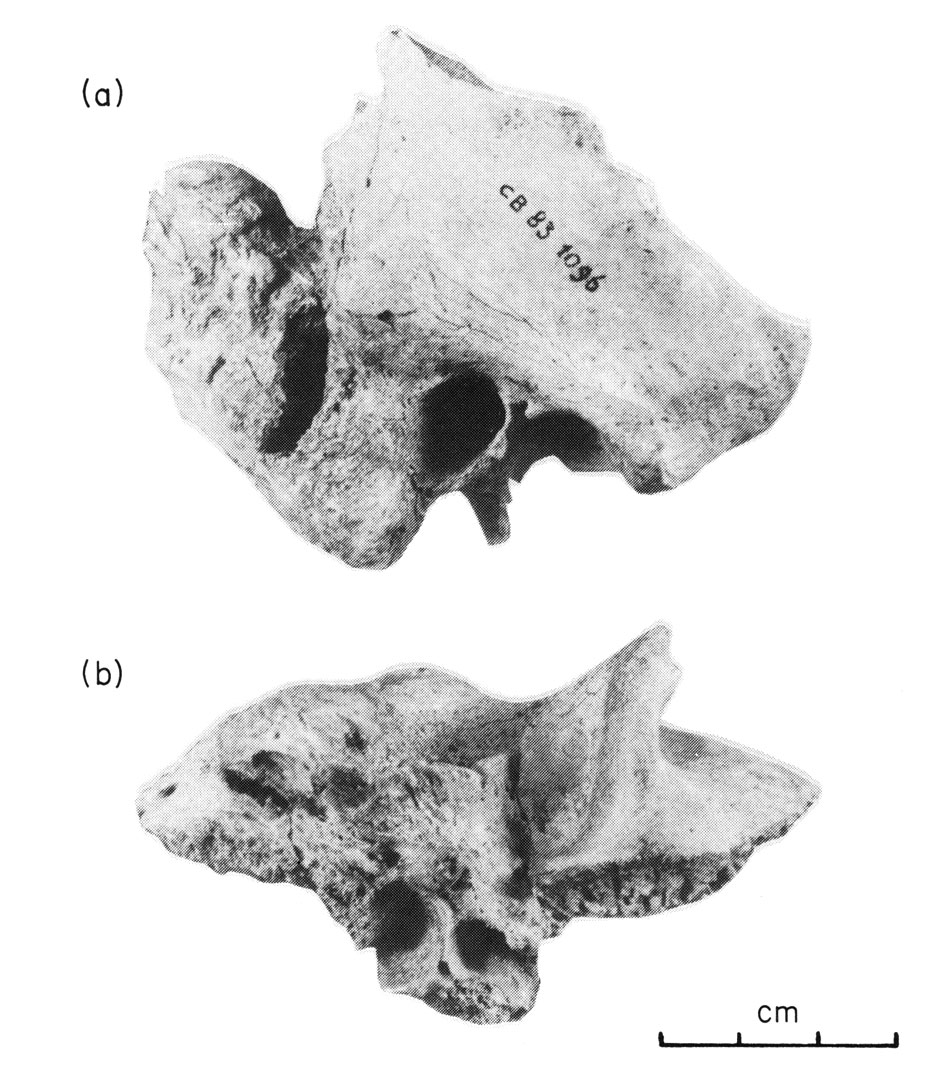

À droite : phalange. Source : Sondaar et coll. 1993
Industries lithiques
Selon Hofmeijer et coll. (1989), l’assemblage lithique de la grotte de Corbeddu n’est pas abondant mais démontre avec succès la méthode de fabrication. La matière première la plus utilisée est une forme locale de calcaire silicifié, du silex et rarement du quartz ou de la goethite. L’industrie la plus ancienne trouvée dans la salle 1 daterait d’environ 14 000 BP (14C). Les auteurs équivalaient l’assemblage lithique de la grotte à l’Épigravettien évolué et tardif de la péninsule.




Environnement

Récemment, une étude anthracologique (analyse des charbons de bois pour déterminer de quelles essences d’arbres ils proviennent) des couches de l’Holocène a été effectuée dans la grotte de Corbeddu (salle 2). Le dépôt semi-continu de la couche la plus basse de ces sédiments composés d’argile fine et de limon sur 4,2 m débuterait il y a 50 000 ans. Les résultats indiquent que presque tous les types de bois pourraient provenir d’espèces végétales actuellement présentes autour de la grotte. Sauf pour deux espèces, le pin noir (Pinus nigra) et le paliure épine du Christ (Paliurus spina-christa). Les auteurs concluent que Paliurus spina-christa était présent au début de l’Holocène comme Pinus nigra, à défaut que celle-ci était une espèce indigène en Sardaigne qui a grandi dans la vallée de Lanaittu. Ces sédiments de l’Holocène montrent également de nombreuses traces de présence humaine et démontre que la grotte était régulièrement visitée au Mésolithique et intensément utilisée au Néolithique (voir Kalis & Schoch, 2019).
Faune

Le cerf de Caziot (Praemegaceros cazioti) daté de 13 590 BP ± 140 ans (14C) a été trouvé dans les gisements du Pléistocène de la grotte du Corbeddu . Il existe des différences de taille entre les cerfs de différentes localités de la Sardaigne, sur la base desquelles une nouvelle espèce de Capo Figari et Santa Lucia a été récemment décrite (Praemegaceros sardus). Le cerf sarde appartient à une lignée qui montre une diminution progressive de sa taille avec le temps. Les fossiles de la grotte de Corbeddu appartiennent à la plus petite taille, typique des dernières populations. Il constitue avec le pika sarde (Prolagus sardus) la majorité des plus de 20 000 espèces fossiles découvertes. Le pika sarde est un animal au taux de reproduction élevé et une masse corporelle d’environ 0,8 kilogramme. Il pourrait avoir été la principale source de nourriture du dhole de Sardaigne et probablement aussi pour les humains du Paléolithique au vu de l’abondance des os brûlés et brisés trouvés dans les couches sédimentaires,

Un squelette presque complet de dhole de Sardaigne (Cynotherium sardous) a été trouvé (salle 2) datant d’environ 11 350 ans ± 100 ans. Ce canidé endémique est connu dans plusieurs localités de Sardaigne et son ancêtre pourrait être l’espèce éteinte Xenocyon. Par la présence de ce petit canidé, la faune insulaire endémique du Pléistocène sarde se distingue des autres faunes insulaires méditerranéennes qui se caractérisent par l’absence de mammifères carnivores autres que les loutres. Le spécimen de cette grotte est le plus petit dhole de Sardaigne et appartient au dernier Pléistocène. Concernant les mammifères trouvés en général on retrouve aussi le hérisson commun (Erinaceus europaeus), un genre de musaraigne (Nesiotites sp.), le loir gris (Glis glis), les rongeurs Microtus (Tyrrhenicola) henseli, Rhagamys orthodon et Apodemus sylvaticus.

L’aigle, animal symbolique ?
Il est intéressant de noter que la plupart des espèces d’oiseaux retrouvés dans la grotte du Corbeddu n’est représentée que par quelques ossements fossiles, sauf pour une espèce qui serait un aigle royal (Aquila chrysaetos) et une espèce de hibou éteinte (Bubo insularis). L’abondance de restes de hibou s’expliquerait simplement en admettant que l’espèce vivait, pour au moins une partie de sa vie, dans la grotte. Mais l’aigle royal, n’a pas pour habitude d’entrer dans les grottes et le plus curieux est le fait que 31 des 37 fossiles trouvés (8 000 à 14 000 ans) de cette espèce sont uniquement des phalanges (malgré la présence d’un fragment de crâne). 28 de ces phalanges ont été trouvées (salle 1) avec la partie distale d’un tibiotarse qui est cassé d’une façon anormale, c’est-à-dire dans une partie très solide entre la diaphyse et l’extrémité distale. Il semblerait que selon cette accumulation et fragmentation, cela soit d’origine anthropique (voir Weesie, 1999).
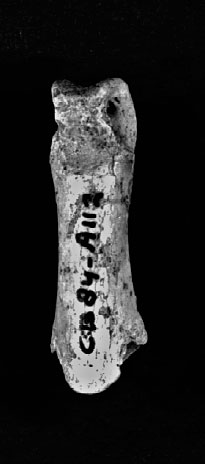

Bien que l’auteur ne mentionne aucune trace de découpes sur ces phalanges et bien qu’elles soient du Paléolithique supérieur, ceci est tout de même intéressant car il existe plusieurs études ultérieures concernant des pratiques symboliques qui seraient ancrées dans les cultures matérielles d’Homo neanderthalensis. Notamment à ce jour, le plus ancien bijou du monde (Krapina, Croatie) daté d’environ 130 000 ans composé de serres d’aigle : elles portent des marques de découpes, de polissages et d’abrasions, suggérant que celles-ci avaient été modelées pour être transformées en bijoux. Il existe un nombre important d’objets présentant des traces de stries sur des serres de rapaces en France et en Italie au Paléolithique moyen. Ceci laisserait suggérer qu’elles auraient été utilisées comme moyen d’expression symbolique par les Néandertaliens dans ces régions.

Un mammouth insulaire
Le mammouth nain endémique de Sardaigne Mammuthus lamarmorai (Major, 1883) serait à ce jour le seul proboscidien connu du Pléistocène sarde. . Il est représenté par des os tarsiens, carpiens et longs, provenant de dépôts éoliens de la glaciation de Riss à Fontana Morimento (Gonnesa) (Acconci 1881). Des empreintes de pas attribuées à cette espèce sont également présente d’après une récente étude d’ichnofossiles (la paléoichnologie est l’analyse des traces d’activité biologique fossilisées). le mammouth nain de Sardaigne atteignait probablement 140-160 cm de hauteur et pesait 500 kg environ.
Grotte de Marras
Au centre de la Sardaigne, une statuette en basalte nommée Vénus de Macomer, haute d’à peu plus de 13 cm, a été découverte en 1949 dans la petite grotte de Marras, sans contrôle scientifique. Il s’agirait d’une thériogyne du Paléolithique supérieur (et non du Néolithique ; voir Mussi, 2012) à tête de pika sarde qui avait une grande part dans l’alimentation de l’homme préhistorique sarde. La forme générale évoquerait le type Gönnersdorf avec une vision latérale préférentielle, des jambes repliées sous des fesses proéminentes, et un petit buste plat, sauf pour un unique sein, sans indication des bras.


Rédaction
Jonathan Ozcelebi
Remerciements à Daniel Zoboli, Paléontologue à l’Université de Cagliari, Sardaigne
Sources :
Bellot-Gourlet, L., C. Mourer-Chauviré, M. Salotti, J. Courtois, G. Poupeau, P. Tramoni, E. Pereira, A. Louchart, C. Oberlin and J. Dubois (2000). « La fin du Pléistocène supérieur et le début de l’Holocène en Corse : Apports paléontologique et archéologique du site de Castiglione (Oletta, Haute-Corse) [The end of the Late Pleistocene and the Early Holocene in Corsica : new paleontological and archaeological data from Castiglione deposit (Oletta, Haute-Corse)]. » Quaternaire: 219-230.
Brown, T., & Walsh, K. (2016). « Holocene Fluvial Dynamics and Geoarchaeology on Mediterranean Islands. In Ghilardi, M. (Ed.), Géoarchéologie des îles de la Méditerranée ». CNRS Éditions : 201-214
Casanovas-Vilar, I., J. A. van Dam, S. Moyà-Solà and L. Rook (2011). « Late Miocene insular mice from the Tusco-Sardinian palaeobioprovince provide new insights on the palaeoecology of the Oreopithecus faunas. » Journal of Human Evolution 61(1): 42- 49.
Germana, F. and C. F. Spoor (1987). « Proportions of the Neolithic Human Maxillae from Sardinia. » Bulletins et Mémoires de la Société d’Anthropologie de Paris: 143-150.
Grifoni, R. and C. Tozzi (2006). « L’émergence des identités culturelles au Paléolithique inférieur : le cas de l’Italie. » Comptes Rendus Palevol 5(1): 137-148.
Hammond, A. S., L. Rook, A. D. Anaya, E. Cioppi, L. Costeur, S. Moyà-Solà and S. Almécija (2020). « Insights into the lower torso in late Miocene hominoid Oreopithecus bambolii. » Proceedings of the National Academy of Sciences 117(1): 278-284.
Hofmeijer, G., C. Alderliesten, K. Van, K. van der Borg, C. Houston, F. Jong, F. Martinit, M. Sangest, P. Sondaar and D. Visser (1989). « Dating of the Upper Pleistocene Lithic Industry of Sardinia. » Radiocarbon 31.
Kalis, A. and W. H. Schoch (2019). « Anthracological studies on the early Holocene sediments of the Grotta di Corbeddu » Nuoro, Sardinia: 193-210
Louchart, A., C. Bedetti and M. Pavia (2005). « A new species of eagle (Aves: Accipitridae) close to the Steppe Eagle, from the Pleistocene of Corsica and Sardinia, France and Italy. » Palaeontographica Abteilung a -Stuttgart- 272: 121-148
Marcus, J. H., C. Posth, H. Ringbauer, L. Lai, R. Skeates, C. Sidore, J. Beckett, A. Furtwängler, A. Olivieri, C. W. K. Chiang, H. Al-Asadi, K. Dey, T. A. Joseph, C.-C. Liu, C. Der Sarkissian, R. Radzevičiūtė, M. Michel, M. G. Gradoli, P. Marongiu, S. Rubino, V. Mazzarello, D. Rovina, A. La Fragola, R. M. Serra, P. Bandiera, R. Bianucci, E. Pompianu, C. Murgia, M. Guirguis, R. P. Orquin, N. Tuross, P. van Dommelen, W. Haak, D. Reich, D. Schlessinger, F. Cucca, J. Krause and J. Novembre (2020). « Genetic history from the Middle Neolithic to present on the Mediterranean island of Sardinia. » Nature Communications 11(1): 939
Mussi, M. (2012). « La Venere di Macomer nel quadro del Pleistocene superiore finale europeo. Atti della XLIV Riunione Scientifica dell’I.I.P.P ». La Preistoria e la Protostoria della Sardegna, 23-28 novembre 2009, vol. II: 383-389. Firenze
Palma di Cesnola, A. (2001). « Le Paléolithique supérieur en Italie ». Collection L’Homme des origines, série Préhistoire d’Europe 9. Éditions Jérôme Millon
Palma di Cesnola, A. (1996). « Le Paléolithique inférieur et moyen en Italie ». Collection L’Homme des origines, série Préhistoire d’Europe 1. Éditions Jérôme Millon
Palombo, M. R., F. Antonioli, V. Lo Presti, M. Mannino, R. Melis, P. Orru, P. Stocchi, S. Talamo, G. Quarta, G. Deiana and S. Altamura (2017). « The late Pleistocene to Holocene palaeogeographic evolution of the Porto Conte area: Clues for a better understanding of human colonization of Sardinia and faunal dynamics during the last 30 ka. » Quaternary International
Palombo, M.R. (2001). « Endemic elephants of the Mediterranean Islands: knowledge, problems and perspectives. » Cavarretta G., Gioia P., Mussi M. & Palombo M.R. (eds), The World of Elephants: Proceedings of the First International Congress, Consiglio Nazionale delle Ricerche, Rome: 486-491
Pillola, G., M. R. Palombo, A. Panarello and D. Zoboli (2020). « The Pleistocene non-hominid vertebrate ichnofossil record of Italy. » Journal of Mediterranean Earth Sciences 12
Sanges, M. (2006). « I primi abitanti della Sardegna. » Darwin Quaderni : Archeologia in Sardegna 1: 30-39
Sondaar, P., R. Elburg, G. K. Hofmeijer, F. Martini, M. Sanges, A. Spaan and H. D. Visser (1995). « The human colonization of Sardinia: a Late-Pleistocene human fossil from Corbeddu cave. »
Sondaar, P. and A. van der Geer (2005). « Evolution and Extinction of Plio-Pleistocene Island Ungulates. » Quaternair 2005: 231-240
Sondaar, P. Y., M. Sanges, T. Kotsakis and P. L. de Boer (1986). « The Pleistocene deer hunter of Sardinia. » Geobios 19(1): 17-31
Spoor, F. (1999). « The human fossils from Corbeddu cave, Sardinia : a reappraisal. In : Reumer, J. W. F & De Vos, J. (eds.) – Elephants have a snorkel ! Papers in honour of Paul Y ; Sondaar. » DEINSEA, 7: 297-302 Spoor, C. F. and P. Y. Sondaar (1986). « Human fossils from the endemic island fauna of Sardinia. » Journal of Human Evolution 15(5): 399-408
van der Geer, A. (2008). Corbeddu Cave and its excavations : 68-92
Weesie, P. D. M. (1999). « Les oiseaux du Pleistocène Supérieur de la grotte de Corbeddu, Sardaigne – in: Reumer, J.W. F. & De Vos, J. (eds.) – Elephants have a snorkel ! Papers in honour of Paul Y. Sondaar. » – DEINSEA 7: 401-409
Zoboli, D., G. Pillola and M. R. Palombo (2018). « The remains of Mammuthus lamarmorai (Major, 1883) housed in the Naturhistorisches Museum of Basel (Switzerland) and the complete “Skeleton-Puzzle”. » Bollettino della Societa Paleontologica Italiana 57
Zoboli, D. and G. Pillola (2018). « New evidences of mammal tracks from the Pleistocene of Gonnesa area (southwestern Sardinia, Italy). »Journal of Mediterranean Earth Sciences 10